- Nach den beiden vorausgehenden – nicht gerade leicht lesbaren – Beiträgen zum Versuch einer Periodisierung der biologischen Evolution anhand eines frei gewählten Komplexitätskriteriums, schälte sich heraus, dass die Besonderheiten des Biologischen sich mit bloßen systemtheoretischen Strukturen nicht fassen lässt. Man kann zwar eine bestimmte Form von Komplexität sichtbar machen, aber das empirisch vorkommende Phänomen des Biologischen kann man auf diese Weise kaum fassen
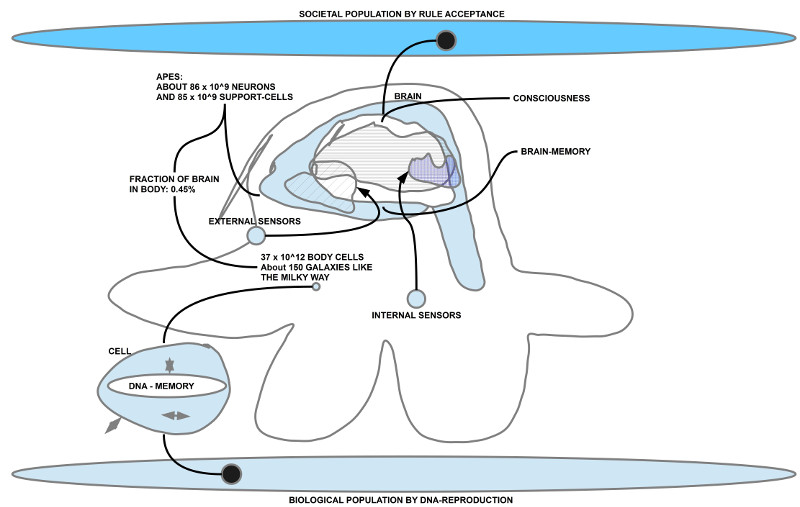
Das Individuum als Bindeglied zwischen der biologischen und der sozialen Dimension - Das Biologische involviert vom Start weg komplexe Kontexte (Erde, Sonnensystem, Galaxie, Universum) und innerhalb des Kontextes Erde sind vom Start weg viele Dimension gleichzeitig wirksam (Eigenschaften der Atome und Moleküle, Umgebungsbeschaffenheiten (Meer, …, Temperaturen, …), Energieverfügbarkeit, chemische Gradienten, Magnetfelder, …). Von Beginn an sind es außerdem komplexe physikalisch-chemische Prozesse, die Prozessumgebungen bereitstellen, innerhalb deren dann eben chemische Prozesse ablaufen können, die zu ihrer eigenen Kodierung und Reproduktion führen. Das ist in vielfacher Hinsicht bemerkenswert, erstaunlich, schwer fassbar, aber, wie sich zeigt, extrem effektiv.
- Die Selbstkodierung von Prozessen inklusive Reproduktionsmechanismen führt zu einer ersten generischen Wissensmaschine (genannt Zelle), die schon auf dieser Ebene das Potential hat, die Menge aller verfügbaren Veränderungen der aktuellen Umgebung zu lernen und kontinuierlich weiter zu entwickeln! Zunächst ist die aktuelle Umgebung – nach heutigem Kenntnisstand – der Ozean; später kam das Land hinzu und dann auch die Luft.
- Die biologische Ära des Lebens war aber gebunden an den Wissensspeicher DNA und jene molekularen Mechanismen, diese zu dekodieren, die dekodierten Informationen chemisch in neue Zellstrukturen zu transformieren. Eine Steigerung der Leistungsfähigkeit war lange nur möglich über die Kooperation von Zellen untereinander. Dies verlangte eine effektive Kommunikation zwischen Zellen. Wenn man weiß, wie schwer sich zwei Menschen tun können, sich selbst über die einfachsten Dinge zu einigen, dann kann man versuchen zu ahnen, was es bedeutet, dass die biologischen Zellen es geschafft haben, im menschlichen Körper ca. 150 Galaxien an Zellen (vom Format der Milchstraße) dazu zu bringen, miteinander im Millisekundenbereich miteinander so zu kooperieren, dass es für Außenstehende aussieht, als ob da ein geschlossenes System auftritt (ein Mensch kann sich so fokussieren, dass er eine Zeitlang nur spielt, oder Musik hört, oder zeichnet oder jemandem zuhört oder Klavier spielt oder …), und während dieser Zeit machen Milliarden, Billionen von Zellen irgendwelche anderen Dinge, aber so, dass diese eine Tätigkeit nicht gestört wird.
- Nach fast 13.8 Mrd Jahren Weltzeit und ca. 3.8 Mrd Jahren biologischer Zeit gab es dann spätestens mit dem homo sapiens eine Lebensform, deren Gehirn die Fähigkeit zum subjektiven Bewusstsein ausgebildet hatte, mittels dem biologische Systeme mit einem Mal auf einer abstrakt-virtuellen Ebene mit anderen biologischen Systemen unabhängig vom DNA-basierten Wissen aktuelle Ereignisse wahrnehmen, abstrahieren, speichern, erinnern, verändern, kombinieren und mittels zeichenbasierten Systemen akut kommunizieren konnten.
- Das eröffnete eine völlig neue Ebene der Wirklichkeitswahrnehmung und deren potentielle Veränderung. Mit einem Mal dauerten Veränderungen nicht mehr viele Generationen der Reproduktion, sondern nun diktierte die Geschwindigkeit des Denkens (Lernen, Planen, …) und kommunikativ vermittelten Kooperierens den Takt möglicher Veränderung.
- Während die moderne Physik den Begriff des ‚Wurmlochs‘ erfunden hat, um eine mögliche Verbindung zwischen zwei Universen beschreiben zu können, ist hier ein neuer Typ von biologischem System in der Lage, von den starren Strukturen des molekularen Wissens durchzutunneln zu den flexiblen Strukturen abstrakt-virtuellen Wissens.
- Die molekulare Maschinerie des Körpers hat sich mit dem Gehirn und darin der Struktur des Bewusstseins eine Möglichkeit geschaffen, molekular-biologisch verankert zu sein und zugleich einen anscheinend ungehemmten Blick in die Gegenwart, die Vergangenheit und die Zukunft aufbauen zu können.
- Die konkrete Empirie eines homo sapiens Körpers zeigt zwar viele konkrete, endliche Rahmenbedingungen im Bereich des Wahrnehmens, Speicherns, Erinnerns, Vorstellens usw., doch die generische Struktur dieser Art des Wissens lässt sich mit technischen Vorrichtungen in viele Richtungen verstärken (künstliche Sensoren und Aktoren, technische Wissensspeicher und Rechenvorrichtungen, Algorithmen zur Fixierung wichtiger Prozesse …). Damit folgt der Revolution durch den homo sapiens als virtuellem Wissensprozessor sehr schnell eine ganze Wolke von technischen Revolutionen, die den Bereich des virtuellen Wissens explosionsartig ausdehnen lassen.
- Hatte der homo sapiens für einen Wimpernschlag der Universalgeschichte die Vision des Individuums als zentraler Bezugsgröße angedacht, so stellt er sich mit seinen eigenen Erfindungen selbst radikal in Frage: der generische virtuelle Wissensraum, der potentiell alles Materielle verändern kann, kennt keine Individuen. Was ist mit dem Individuum von Typ eines homo sapiens? War dies nur ein extrem kurzer Traum oder steht das biologische Leben vor seiner nächsten großen Entdeckung? Alle bekannten sogenannten Religionen sind aus Sicht des modernen Wissens ‚Wissensschrott‘; sie stimmen vorne und hinten nicht; war es das mit einem potentiellen Schöpfer-Gott oder haben wir wichtige Dinge einfach noch nicht gut genug verstanden?
- Da wir fast alles noch kaum verstehen, nur ansatzweise, fragmentarisch, müssen wir bereit sein, dass neue Erkenntnisse alles, was wir bisher kennen, nochmals komplett auf den Kopf zu stellen oder von innen nach außen drehen oder …. Das Tröstliche an allem ist, wir sind keine ‚Fremden‘ in diesem Prozess; wir alle sind Teil dieses unfassbaren Lebensprozesses, wir sind ‚ins Spiel gebracht worden‘, wir erleben es, können handeln auch wenn es so unfassbar unbedeutend erscheinen mag. Selbst die größten Diktatoren einer Zeit oder die größten egoistischen Kapitalisten haben keine wirkliche Chance, wenn sie gegen den Prozess arbeiten. Bildung mit wahrem Wissen und Kommunikation sind die zentralen Treiber des Lebens. Die Zukunft beginnt immer jetzt, in diesem Moment. Jetzt entscheiden wir, wo wir morgen stehen werden. Wer nach hinten schaut oder bewahren will, was jetzt ist, hat schon verloren.
Einen Überblick über alle Blogeinträge von Autor cagent nach Titeln findet sich HIER.
Einen Überblick über alle Themenbereiche des Blogs findet sich HIER.
