Journal: Philosophie Jetzt – Menschenbild, ISSN 2365-5062, 27.August 2017
URL: cognitiveagent.org
Email: info@cognitiveagent.org
Autor: Gerd Doeben-Henisch
Email: gerd@doeben-henisch.de
Letzte Altualisierung: 27.Aug.2017 - 17:37h Es gibt eine Weiterentwicklung dieses Beitrags in einem Folgebeitrag!
Überblick
Eingeleitet durch wissenschaftsphilosophische
Überlegungen wird versucht, die Entwicklung der
Säugetiere bis hin zum homo sapiens anhand der aktuellen
Forschungsdaten abzubilden. Das Hauptaugenmerk liegt
auf der allgemeinen Struktur. Für die vielen Details sei auf
die Literatur verwiesen, die angegeben wird.
I. KONTEXT
Eine der Leitfragen dieses Blogs ist die Frage nach
dem neuen Menschenbild, speziell auch im Kontext
der Diskussion um die Zukunft von Menschen und
intelligenten Maschine.
Wer nach der Zukunft des Menschen fragt,
braucht ein gutes Bild vom aktuellen Menschen und
seiner Entstehungsgeschichte, um auf dieser Basis
Überlegungen zu einer möglichen Zukunft anstellen zu
können.
Während zur biologischen Evolution allgemein schon
einige Blogbeiträge geschrieben wurden, fehlt es im Blog
an konkreten Daten zur Entwicklung unmittelbar vor dem
Auftreten des homo sapiens, etwa in dem Zeitfenster -10
Mio Jahren bis ca. -30.000 Jahren vor dem Jahr 0. Dies
soll hier in einem ersten Beitrag nachgeholt werden.
II. WISSENSCHAFTLICHE SICHTWEISEN
Bei der Frage nach der Entwicklung des homo
sapiens spielen mehrere wissenschaftliche Disziplinen
ineinander. Einmal ist es die Geologie (Siehe: [WD17g]),
die den Kontext Erde untersucht; dann die Klimatologie
(Siehe: [WD17n]), die sich mit den klimatischen
Verhältnissen im Kontext Erde beschäftigt. Für das
Phänomen des Lebens selbst ist die Biologie zuständig,
speziell die Evolutionsbiologie (Siehe: [SWW13],
[WD17e]). Als Teil der Evolutionsbiologie sind noch
zu nennen die Molekularbiologie (Siehe: [WD17s]) mit
der Genetik (Siehe: [WD17f]). Ferner könnte man als
Teil der Evolutionsbiologie auch noch die Paläontologie
(Siehe: [WD17u], [Par15]) nennen und auch die
Archäologie (Siehe: [WD17a]). Wobei das Wechselspiel
von Evolutionsbiologie und Archäologie nicht ganz
so klar erscheint. Viele weitere wissenschaftliche
Disziplinen tauchen innerhalb der genannten Disziplinen
in unterschiedlichsten Kontexten auf.
Diese Vielfalt spiegelt ein wenig die Komplexität der
Phänomene wieder, um die es hier geht. Der Autor
cagent selbst betrachtet die hier zu verhandelnden
empirischen Phänomene aus Sicht der Philosophie
mit den Schwerpunkten Erkenntnisphilosophie, die
Überschneidungen hat mit Phänomenen wie z.B.
‚Lernen’ und ’Intelligenz’. Dies sind Themen, die
feste Orte auch in der Psychologie haben, heute
oft parallelisiert mit der Gehirnforschung (letztere
methodisch gesehen ein Teil der Biologie).
Angesichts dieser Komplexität ist es praktisch
unmöglich, ein völlig konsistentes, einheitliches Bild der
Phänomene zu zeichnen. An dieser Stelle ist es das
Anliegen von cagent, einen ’hinreichenden’ Überblick
über die ’unmittelbare’ Vorgeschichte des homo sapiens
zu bekommen.
Der homo sapiens ist jene biologische Art (Spezies),
die zur Gattung homo gerechnet wird, die sich aus
dem biologischen Formenstrom über Jahrmillionen
herausgeschält hat. Es zeigt sich, dass die zeitliche
Abgrenzung, wann genau ’das Menschliche’ anfängt,
und wann das ’Tierische’ aufhört, irgendwie beliebig
erscheint. Der homo sapiens ab ca. -30.000 besitzt
natürlich Eigenschaften, die wir beschreiben können
wie Körperbau, genetisch bestimmte Erbanlagen,
typische Verhaltensweisen, aber diese Eigenschaften
tauchen nicht abrupt in der Geschichte auf, sind nicht
irgendwann einfach so da, sondern man findet in
der vorausgehenden Zeit eine große Formenvielfalt
in den Artefakten, mit unterschiedlichen genetischen
Distanzen zum heutigen homo sapiens. Daraus muss
man schließen, dass es einen viele Millionen dauernden
Prozess des Formenwandels gab, innerlich genetisch
und äußerlich durch die jeweiligen geologischen und
klimatologischen Verhältnisse beeinflusst, die sich
zudem noch verknüpfen mit der jeweiligen Pflanzen- und
Tierwelt. Alle diese genannten Faktoren waren selbst
einem kontinuierlichen Wandel unterworfen.
Wenn die Grenzziehung zwischen dem ’Tierischen’
und dem ’Menschlichen’ von daher nicht ganz scharf
gezogen werden kann, ist auch eine Zeitangabe dazu,
wie weit zurück in der Zeit man gehen soll, um die
Vorgeschichte’ zu beschreiben, relativ, d.h. abhängig
von den Kriterien, die man bei der Analyse anlegen
will.
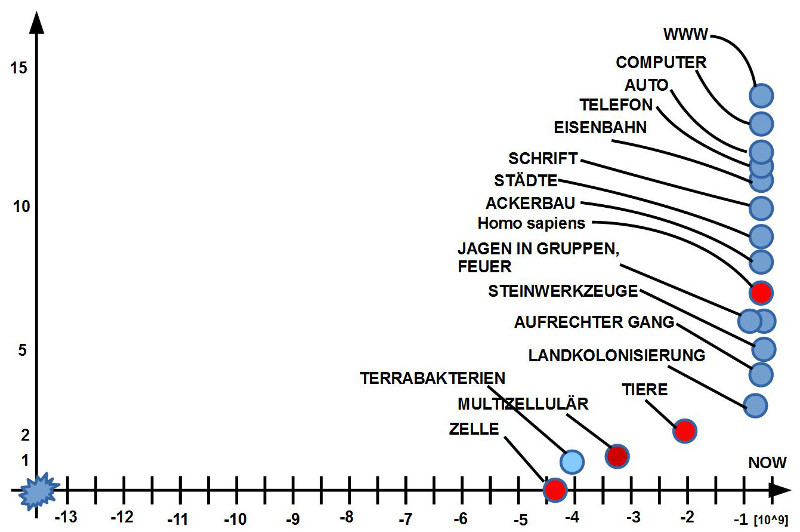
In diesem Beitrag wurde der Startpunkt für die
Beschreibung bei den Lebensformen gewählt, die die
Biologen ’Primaten’ nennen, und zwar spezieller den
Punkt der Aufspaltung in die Strepsirrhini und Haplorhini
(Siehe: [WE17l] und [WD17r]), die sich um etwa -80
Mio Jahren ereignet haben soll. Aus Sicht der heutigen
menschlichen Geschichte, wo 100 Jahre oder gar 1000
Jahre eine lange Zeit sind, wirken diese 80 Millionen
Jahre sehr, sehr lang. Innerhalb der Geschichte des
Lebens mit ca. 3.5 Milliarden Jahre fallen die 0.08 Mrd
Jahre seit dieser Aufspaltung nahezu kaum ins Gewicht,
es sind gerade mal 2.2% der gesamten Entwicklungszeit
des biologischen Lebens. Betrachtet man dagegen nur
die Zeit seit dem Auftreten der Lebensform homo, die
dem heute bekannten Menschlichem schon nahe kommt
(etwa ab -2.5 Mio), dann schrumpft der Zeitanteil auf
0.071 % der Entwicklungszeit des biologischen Lebens
zusammen. Umgerechnet auf das 12-Stunden Ziffernblatt
einer Uhr mit 720 Minuten würde die Entstehung der
Lebensform homo die letzte halbe Minute auf dem
Ziffernblatt ausmachen. Die Lebensform homo sapiens,
zu der wir uns zählen, tauchte frühestens um -190.000 in
Afrika auf. Das wären auf dem Ziffernblatt dann (bei ca.
81.000 Jahren pro Sekunde) die letzten 2.3 Sekunden.
Im Spiel des Lebens erscheint dies nicht viel. Betrachtet
man aber, was sich allein in den letzten ca. 10.000
Jahren ereignet hat, und hier speziell nochmals in den
letzten 100 Jahren, dann legt sich der Schluss nahe,
dass die Lebensform homo sapiens offensichtlich über
Fähigkeiten verfügt, die gegenüber der Vorgeschichte
von ca. 3.5 Mrd Jahren etwas qualitativ ganz Neues
sichtbar macht. Autor cagent ist sich nicht sicher,
ob der homo sapiens selbst bislang wirklich begreift,
was hier passiert, welche Rolle er in diesem Prozess
spielt. Auf der einen Seite zeichnet sich eine immer
größere Zerstörungskraft ab (die auch stattfindet), auf
der anderen Seite deuten sich konstruktive Potentiale
an, die alles übersteigen, was bislang vorstellbar war.
III. DEUTUNGEN: MATERIAL, MUSTER, FUNKTION, KONTEXTE
Die Tätigkeit der eingangs erwähnten Wissenschaft
kann man verstehen als eine Deutung, ausgeführt in
einem Deutungsprozess. Diese Deutung beginnt bei der
Bestimmung der Substanzen/ Materialien, die Forscher
vorfinden. Ist das eine Gesteinsart, sind das Knochen,
sind das pflanzliche Bestandteile …. ? Ein anderer
Aspekt ist die Frage nach ’Formen’ und ’Mustern’:
kann man an dem Material auffällige Formen oder
Muster erkennen, dann auch im Vergleich zu anderen
Materialien? Schließlich auch die Frage nach möglichen funktionalen Zusammenhängen’: wenn es ein Knochen
ist, in welchem Zusammenhang eines Knochengerüsts
kommt er vor? Wenn etwas ein Zahn sein soll, wie sah
das zugehörige Gebiss aus? Oder ist dieser Knochen Teil
eines Werkzeugs, einer zu unterstellenden Handlung,
die das Stück benutzt hat? Schließlich, in welchem
Kontext kommt ein Material vor? Ist es zufälliger Kontext,
ein Kontext durch einen geologischen Prozess, ein
Kontext erzeugt durch Verhalten von Lebewesen?
Schon bei diesen Fragen bieten sich eine Vielzahl von
Deutungsmöglichkeiten, bestehen viele Ungewissheiten.
IV. DEUTUNGEN 2: ZEITLICHE ABFOLGE
Was Forscher zur Evolutionsbiologie besonders
interessiert, ist das Erfassen von zeitlichen Abfolgen:
unter Voraussetzung eines bestimmten Zeitmaßes
möchte die Evolutionsbiologie wissen, ob ein
Gegenstand/ Artefakt A im Sinne des Zeitmaßes ‚vor’ oder ’nach’ einem anderen Gegenstand/ Artefakt B ‚anzuordnen’ ist.
Diese Frage macht nur Sinn, wenn man neben
einem definierten Zeitmaß auch annehmen darf
(muss), dass sich die Erde als Generalumgebung aller
vorfindbaren Materialien/ Artefakte grundsätzlich in
einem Veränderungsmodus befindet, dass also die
Erde zu zwei verschiedenen Zeitpunkten grundsätzlich
verschieden sein kann.
Dass sich am Kontext Erde Veränderungen feststellen
lassen, dies haben Menschen schon sehr früh erleben
können: Temperatur, Regen oder nicht Regen, Tag und
Nacht, Wachstum der Pflanzen, Geboren werden und
Sterben, usw. Es passierte aber erst im 17.Jahrhundert,
dass die Fragestellung nach dem Vorher und Nachher in
der Entwicklung der Gesteine mit Nils Stensen (nicolaus
steno) eine systematische Form fand, aus der sich nach
und nach die moderne Geologie entwickelte (Siehe:
[WD17h]).
Erst durch die wissenschaftliche Geologie wissen
wir zunehmend, dass die Erde selbst ein dynamisches
System ist, das sich beständig verändert, wo sich
ganze Kontinente bilden, verschieben, verformen; wo
Vulkanismus stattfindet, Erosion, Klimaänderungen, und
vieles mehr. Erst durch die geologische Sehweise konnte
man nicht nur verschiedene Zustände der Erde entlang
einem definierten Zeitmaß identifizieren, sondern damit
dann auch Veränderungen in der Zeit’ sichtbar machen.
Dieses geologische Wissen vorausgesetzt, besteht
plötzlich die Möglichkeit, ein Material/ Artefakt einer
erdgeschichtlichen Phase, einem Zeitpunkt in einem
Veränderungsprozess, zuzuordnen. Die Geologie hat – mittlerweile unterstützt durch viele Spezialgebiete, wie z.B. auch die Klimatologie (Siehe:
[WD17n]) – unter anderem eine zeitliche Abfolge von
Vulkanausbrüchen in den verschiedenen Regionen
der Erde identifizieren können und auch das sich
verändernde Klima.
So spricht man in der Klimatologie von sogenannten
Eiszeitaltern’ (Siehe: [WD17d]). In der schwachen
Version einer Definition von Eiszeitalter geht man davon
aus, dass mindestens eine Polkappe vereist ist. Die
letzte Eiszeit dieser Art fand statt um -33.5 Mio Jahren.
In der starken Version geht man davon aus, dass beide
Polkappen vereist sind. Die letzte Eiszeit dieser Art
begann um ca. -2.7 Mio Jahren und hält bis heute
an. In dieser Zeit gab es unterschiedliche Kalt- und
Warm-Phasen. Seit ca. -1 Mio Jahren haben sich 6
mal Kaltzeiten wiederholt: ca. -0.9 Mio, -0.77 Mio, -0.6
Mio, -0.48 Mio, -0.35 Mio, -12.000 (siehe: [WD17o],
[Rot00]:SS.173ff ).
Ein anderer starker Faktor, der das Klima
beeinflussen kann, sind Supervulkanausbrüche
(Siehe: [WD17w]). Hier eine Zusammenstellung
von Eiszeitaltern mit Kaltphasen in Kombination mit
den Supervulkanausbrüchen sofern sie das frühe
Ausbreitungsgebiet von homo und homo sapiens berührt
haben (wobei auch andere große Ausbrüche sich
weltweit auswirken konnten)(man beachte, dass die
Zeitangaben mit großen Unschärfen versehen sind):
- Eiszeit: ab ca. -2.7 Mio Jahren
- Vulkan:-1 Mio Äthiopien
- Vulkan: -788.000 Indonesien
- Kaltzeit: ca. -0.77 Mio Jahren
- Kaltzeit: ca. -0.6 Mio Jahren
- Vulkan: -500.000 (+/- 60.000) Äthiopien
- Kaltzeit: ca. -0.48 Mio Jahren
- Vulkan: -374.000 Italien
- Kaltzeit: ca. -0.35 Mio Jahren
- Vulkan:-161.000 Griechenland
- Vulkan: -74.000 Indonesien
- Vulkan:-50.000 Italien
- Vulkan:-39.000 Italien
- Kaltzeit: ca. -12.000
Bei der Entstehung von Eiszeiten spielen eine Vielzahl
von Faktoren eine Rolle, die ineinandergreifen. Sofern
es sich um periodische Faktoren handelt, kann sich dies
auf den Periodencharakter von Kalt- und Warmzeiten
auswirken (siehe: [WD17o], [Rot00]:SS.173ff ). Die globale Erwärmung, die
aktuell beklagt wird, ist ein Ereignis innerhalb eines noch
existierenden Eiszeitalters. Insofern ist die Erwärmung
eigentlich keine Anomalie, sondern eher die Rückkehr
zum ’Normalzustand’ ohne Eiszeitalter. Wobei sich
natürlich die Frage stellt, welcher Zustand der Erde ist
eigentlich ’normal’? Kosmologisch betrachtet – und darin
eingebettet die Wissenschaften von der Erde – wird
die Erde in einem Zustand enden, der nach heutigem
Wissen absolut lebensfeindlich sein wird (siehe: [WD17p],
[WE17b], [WE17c]). Für die Erde ist dieser Zustand
normal’, weil es dem physikalischen Gang der Dinge
entspricht, aus Sicht der biologischen Lebensformen
ist dies natürlich überhaupt nicht ’normal’, es ist ganz
und gar ’fatal’.
Insofern wird hier schon deutlich, dass
die innere Logik des Phänomens ‚biologisches Leben‘
nicht automatisch kongruent ist mit einem aktuellen
Lebensraum. Das Phänomen des biologischen Lebens
manifestiert einen Anspruch auf Geltung, für den
es im Licht der physikalischen Kosmologie keinen
natürlichen’ Ort gibt. Das biologische Leben erscheint
von daher als eine Art ’Widerspruch’ zum bekannten
physikalischen Universum, obgleich es das physikalische
Universum ist, das das biologische Leben mit ermöglicht.
V. DEUTUNGEN 3: ENTWICKLUNG VON KOMPLEXITÄT
Wenn man so weit vorgestoßen ist, dass man
Materialien/ Artefakte auf einer Zeitachse anordnen kann,
dann kann man auch der Frage nachgehen, welche
möglichen Veränderungen sich entlang solch einer
Zeitachse beobachten lassen: Bleibt alles Gleich? Gibt
es Änderungen? Wie lassen sich diese Veränderungen
klassifizieren: werden die beobachtbaren Phänomene
einfacher’ oder ’komplexer’?
Um solche eine Klassifikation in ’einfach’ oder
komplex’ vorzunehmen, braucht man klare Kriterien für
diese Begriffe. Aktuell gibt es aber keine einheitliche, in
allen Disziplinen akzeptierte Definition von ’Komplexität’.
In der Informatik wird ein Phänomen relativ zu
einem vorausgesetzten Begriff eines ’Automaten’ als
komplex’ charakterisiert: je nachdem wie viel Zeit
solch ein Automat zur Berechnung eines Phänomens
benötigt oder wie viel Speicherplatz, wird ein Phänomen
als mehr oder weniger ’komplex’ eingestuft (Siehe
dazu: [GJ79]). Dieser vorausgesetzte Automat ist eine
sogenannte ’Turingmaschine’. Dieses Konzept entstand
in der Grundlagendiskussion der modernen Mathematik
um die Wende vom 19. zum 20.Jahrhundert, als sich
die Mathematiker (und Logiker) darüber stritten, unter
welchen Bedingungen ein mathematischer Beweis
für einen Menschen (!) als ’nachvollziehbar’ gelten
kann. Nach gut 30 Jahren heftigster Diskussionen fand
man mehrere mathematische Konzepte, die sich als
äquivalent erwiesen. Eines davon ist das Konzept der
Turingmaschine, und dieses gilt als das ’einfachste’
Konzept von allen, das sich seit 1936/7 bisher in
allen Widerlegungsversuchen als stabil erwiesen hat.
Dies ist zwar selbst kein unwiderleglicher logischer
Beweis, aber ein empirisches Faktum, was alle Experten
bislang glauben lässt, dass mit diesem Konzept eine
zentrale Eigenschaft des menschlichen Denkens
eine konzeptuelle Entsprechung gefunden hat, die
sich formal und empirische experimentell überprüfen
lässt. So, wie die Physiker zum Messen Standards
entwickelt haben wie das ’Kilogramm’, das ’Meter’
oder die ’Sekunde’, so haben die Informatiker zum
Messen der ’Komplexität’ eines Phänomens relativ zur
(menschlichen) Erkenntnisfähigkeit die ’Turingmaschine’
(samt all ihren äquivalenten Konzepten) gefunden. Der
Vorteil dieser Definition von Komplexität ist, dass man
über das zu klassifizierende Phänomen vorab nahezu
nichts wissen muss. Darüber hinaus macht es Sinn, das
menschliche Erkennen als Bezugspunkt zu wählen, da
die Frage der Komplexität jenseits des menschlichen
Erkennens keinen wirklichen Ort hat.
Zurück zum Ausgangspunkt, ob sich im ’Gang der
Dinge’ auf der Erde Phänomene erkennen lassen,
die ’im Lauf der Zeit’ an Komplexität zunehmen,
deutet sich Folgendes an: es scheint unbestritten,
dass die Beschreibung einer biologischen ’Zelle’
(siehe: [AJL+15]) einen erheblich größeren Aufwand
bedeutet als die Beschreibung eines einzelnen Moleküls.
Zellen bestehen aus Milliarden von Molekülen, die
in vielfältigsten funktionellen Zusammenhängen
miteinander wechselwirken. Der Übergang von einzelnen
Molekülen zu biologischen Zellen erscheint von daher
gewaltig, und es ist sicher kein Zufall, dass es bis heute
kein allgemein akzeptiertes Modell gibt, das diesen
Übergang vollständig und befriedigend beschreiben
kann.
Für den weiteren Verlauf der biologischen Evolution
gibt es zahllose Phänomene, bei denen eine Vielzahl
von Faktoren darauf hindeuten, dass es sich um eine
Zunahme von Komplexität’ im Vergleich zu einer
einzelnen Zelle handelt, wenngleich manche dieser
Komplexitäts-Zunahmen’ Milliarden oder hunderte von
Millionen Jahre gebraucht haben. Im Fall der Entwicklung
zum homo sapiens ab ca. -80 Millionen Jahre gibt es
auch solche Phänomene, die sich aber immer weniger
nur alleine im Substrat selbst, also im Körperbau
und im Gehirnbau, festmachen lassen, sondern wo
das ’Verhalten’ der Lebewesen ein Indikator ist für
immer komplexere Wechselwirkungen zwischen den
Lebewesen und ihrer Umwelt.
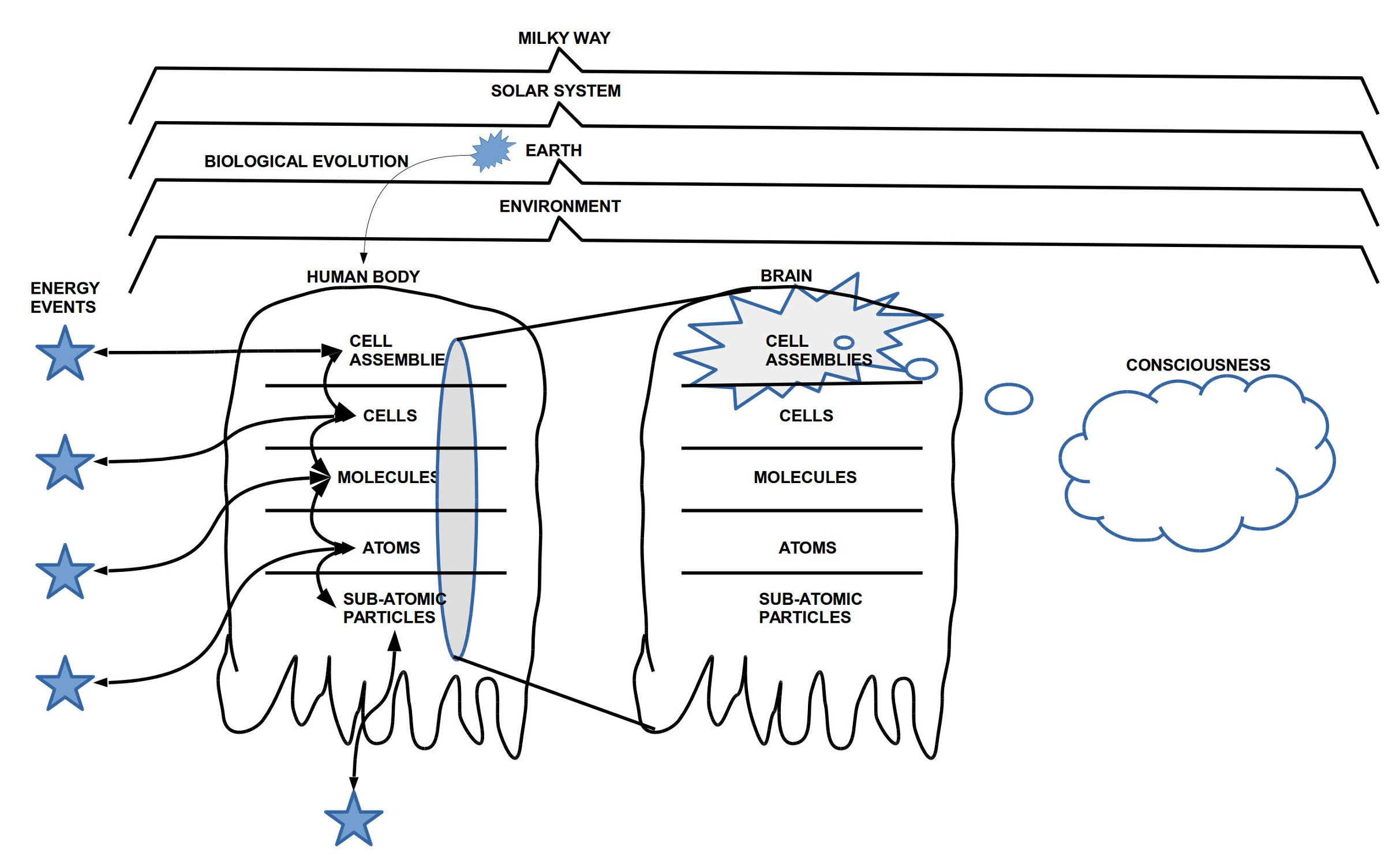
Der Körper des homo sapiens selbst umfasst ca.
37 Billionen (10^12) Körperzellen, dazu im Innern des
Körpers geschätzte ca. 100 Billionen Bakterien, und
zusätzlich auf der Körperoberfläche ca. 224 Milliarden
Bakterien (siehe dazu [Keg15]). Diese ca. 137 Billionen
Zellen entsprechen etwa 437 Galaxien im Format
der Milchstraße. Während Menschen beim Anblick
des Sternenhimmels zum Staunen neigen, bis hin
zu einer gewissen Ergriffenheit über die Größe (und
Schönheit) dieses Phänomens, nehmen wir einen
anderen menschlichen Körper kaum noch wahr (falls
er sich nicht irgendwie auffällig ’inszeniert’). Dabei
ist der menschliche Körper nicht nur 437 mal größer in seiner Komplexität
als die Milchstraße, sondern jede einzelne Zelle ist
ein autonomes Individuum, das mit den anderen auf
vielfältigste Weise interagiert und kommuniziert. So kann
eine einzelne Gehirnzelle bis zu 100.000 Verbindungen
zu anderen Zellen haben. Körperzellen können über
elektrische oder chemische Signale mit vielen Milliarden
anderer Zellen kommunizieren und sie beeinflussen.
Bakterien im Darm können über chemische Prozesse
Teile des Gehirns beeinflussen, das wiederum aufgrund dieser Signale Teile des
Körpers beeinflusst. Und vieles mehr. Obgleich
die Erfolge der modernen Wissenschaften in den letzten
20 Jahren geradezu atemberaubend waren, stehen wir
in der Erkenntnis der Komplexität des menschlichen
Körpers noch weitgehend am Anfang. Niemand hat
bislang eine umfassende, zusammenhängende Theorie.
Dazu kommen noch die vielen immer komplexer
werden Muster, die sich aus dem Verhalten von
Menschen (und der Natur) ergeben. Zusätzlich wird das Ganze
stark beeinflusst von modernen Technologi wie z.B. der
Digitalisierung.
VI. DEUTUNGEN4: SELBSTREFERENZ: CHANCE UND
RISIKO
Ist man also zur Erkenntnis einer Zunahme an
Komplexität vorgestoßen, gerät das Erkennen vermehrt
in einen gefährlichen Zustand. Das Erkennen von
Zunahmen an Komplexität setzt – nach heutigem
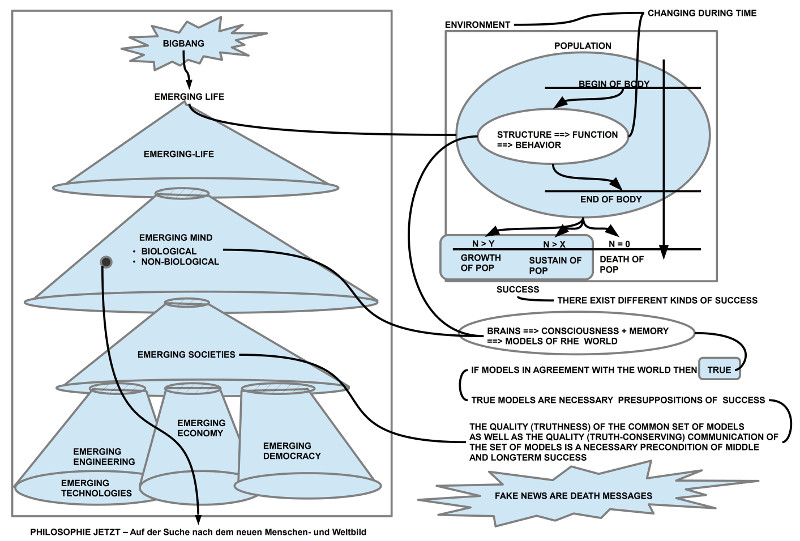
Wissensstand – symbolisch repräsentierte ’Modelle’
voraus, ’Theorien’, mittels deren das menschliche
(und auch das maschinelle) Denken Eigenschaften
und deren Anordnung samt möglichen Veränderungen
repräsentieren’. Sobald ein solches Modell vorliegt, kann
man damit die beobachteten Phänomene ’klassifizieren’
und in ’Abfolgen’ einordnen. Die ’Übereinstimmung’
von Modell und Phänomen erzeugt psychologisch ein
befriedigendes’ Gefühl. Und praktisch ergibt sich daraus
meist die Möglichkeit, zu ’planen’ und Zustände ’voraus
zu sagen’.
Je komplexer solche Modelle werden, um so größer
ist aber auch die Gefahr, dass man nicht mehr so leicht
erkennen kann, wann diese Modelle ’falsch’ sind. Solche
Modelle stellen Zusammenhänge (im Kopf oder in der
Maschine) her, die dann vom Kopf in die Wirklichkeit
außerhalb des Körpers ’hinein gesehen’ werden, und
mögliche Alternativen oder kleine Abweichungen können
nicht mehr so ohne weiteres wahrgenommen werden.
Dann hat sich in den Köpfen der Menschen ein bestimmtes
Bild der Wirklichkeit ’festgesetzt’, das auf Dauer
fatale Folgen haben kann. In der Geschichte der empirischen
Wissenschaften kann man diese Prozesse mit
zahlreichen Beispielen nachvollziehen (siehe den Klassiker:
[Kuh62]). Dies bedeutet, je umfassender Modelle
des Erkennens werden, um so schwieriger wird es auf
Dauer – zumindest für das aktuelle menschliche Gehirn
das ’Zutreffen’ oder ’Nicht-Zutreffen’ dieser Modelle
zu kontrollieren.
Nachdem mit dem Gödelschen ’Unentscheidbarkeitstheorem’
schon Grenzen des mathematischen Beweisens sichtbar wurden (Siehe: [WD17q]),
was dann mit der Heisenbergschen ’Unschärferelation’
(Siehe: [WD17j]) auf das empirischen Messen erweitert
wurde, kann es sein, dass das aktuelle menschliche
Gehirn eine natürliche Schranke für die Komplexität
möglicher Erklärungsmodelle bereit hält, die unserem
aktuellen Erkennen Grenzen setzt (Grenzen des Erkennens
werden im Alltag in der Regel schon weit vorher
durch psychologische und andere Besonderheiten des
Menschen geschaffen).
VII. PERIODISIERUNGEN: BIS HOMO SAPIENS
Wie schon angedeutet, ist das Vornehmen einer
Periodisierung ein Stück willkürlich. Autor cagent hat
den Zeitpunkt der Aufspaltung der Primaten um etwa
-80 Mio Jahren vor dem Jahr 0 gewählt. Dabei gilt
generell, dass solche Zeitangaben nur Näherungen sind,
da die zugehörigen Wandlungsprozesse sich immer als
Prozess über viele Millionen Jahre erstrecken (später
dann allerdings immer schneller).
Bei der Datierung von Artefakten (primär
Knochenfunden, dazu dann alle weiteren Faktoren,
die helfen können, den zeitlichen Kontext zu fixieren),
gibt es einmal den Ansatzpunkt über die äußere und
materielle Beschaffenheit der Artefakte, dann aber
auch – im Falle biologischer Lebensformen – den
Ansatzpunkt über die genetischen Strukturen und
deren Umformungslogik. Über die Genetik kann man
Ähnlichkeiten (Distanzen in einem Merkmalsraum)
zwischen Erbanlagen feststellen sowie eine ungefähre
Zeit berechnen, wie lange es gebraucht hat, um von
einer Erbanlage über genetische Umformungen zu
einer anderen Erbanlage zu kommen. Diese genetisch
basierten Modellrechnungen zu angenommenen Zeiten
sind noch nicht sehr genau, können aber helfen,
die Materie- und Formen-basierten Zeitangaben zu
ergänzen.
- Ordnung: Primates (Siehe: [SWW13]:Kap.5.2)
(Aufteilung ab ca. -80 Mio) –->Strepsirrhini (Lorisi-,
Chiromyi-, Lemuriformes) und Haplorhini (Tarsier,
Neu- und Altweltaffen (einschließlich Menschen))
(Siehe: [SWW13]:S.428,S.432, S.435 [WE17l],
[WD17r]) - Unterordnung: Haplorrhini (Aufteilung ab ca. -60
Mio) (Siehe: [WE17l]) –->Tarsiiformes und Simiiformes
Nebenordnung: Simiiformes (Aufteilung ab ca. –
42.6 Mio) -–>Platyrrhini (Neuwelt- oder Breitnasenaffen)
und Catarrhini (Altwelt- oder Schmalnasenaffen)
(Siehe: Siehe: [SWW13]:S.428, [WE17l]) - Teilordnung: Catarrhini (Altwelt- oder Schmalnasenaffen)
(Aufteilung ab ca. -29/-25 Mio) -–>Cercopithecoidea
(Geschwänzte Altweltaffen) und Hominoidea
(Menschenartige) (Siehe: Siehe: [WE17l] und
[WD17r])- Überfamilie: Hominoidea (Menschenartige)
(Aufteilung ab ca. -20 Mio/ -15 Mio) –>Hylobatidae
(Gibbons)und Hominidae (Große Menschenaffen
und Menschen) (Siehe: [WD17r]) - Aufspaltung der Menschenaffen (Hominidae) in die
asiatische und afrikanische Linie (ca. -11 Mio)
(Siehe: [WD17r])- Familie: Hominidae (Menschenaffen)(Aufteilung ab
ca. -15Mio/-13 Mio in Afrika) –>Ponginae (Orang-
Utans) und Homininae (Siehe: [WD17r])- Unterfamilie: Homininae
Aufteilung der Homininae (ab ca. -9 Mio/ -8 Mio) –>
Tribus: Gorillini und Hominini (Siehe: [WE17d])- Gattung: Graecopithecus (Süden von Griechenland)
- Spezies/ Art: Graecopithecus freybergi (Siehe: [WD17i]) (ca. -7.2 Mio)
- Gattung: Sahelanthropus (ab ca. -7.0/ -6.0 Mio)
- Spezies/ Art: Sahelanthropus tchadensis
(Siehe: [WD17v] [WE17k]) (im Tschad)
- Tribus (Stamm/ Tribe): Hominini
- Aufteilung der Hominini (ab ca. -6.6/-4.2 Mio)
(Siehe: [SWW13]:S.435, [WE17d]) -–>Pan
(Schimpansen) und Homo (Die Lebensform
Panina bildet einen Unterstamm zum
Stamm ’homini’. Für die Trennung zwischen
Schimpansen (Pan) und Menschen (Homo) wird
ein komplexer Trennungsprozess angenommen,
der viele Millionen Jahre gedauert hat. Aktuelle
Schätzungen variieren zwischen ca. -12 Mio und
-6-8 Mio Jahren (Siehe: [WE17a])- Gattung: Orrorin tugenensis (ab ca. -6.2 bis
ca. -5.65 Mio) (Siehe: [WD17t]) - Gattung: Ardipithecus (ab ca. -5.7 Mio bis ca.
-4.4 Mio) (Siehe: [WD17b]) - Gattung: Australopithecus anamensis (ab
ca. -4.2 Mio bis ca. -3.9 Mio) (Siehe:
[SWW13]:S.475f) - Gattung: Australopithecus (ab ca. -4 Mio bis
ca. -2/-1.4 Mio) (Siehe: [SWW13]:S.475f) - Gattung: Australopithecus afarensis (ab
ca. -3.5 Mio bis ca. -3 Mio) (Siehe:
[SWW13]:S.476) - Gattung: Kenyanthropus platyops (ab ca. –
3.5/ -3.3 Mio) (Siehe: [WD17m]) Kann
möglicherweise auch dem Australopithecus
zugerechnet werden (Siehe: [SWW13]:S.475,
479). - Gattung: Australopithecus africanus (ab
ca. -3.2 Mio bis ca. -2.5 Mio) (Siehe:
[SWW13]:S.477) - Gattung: Australopithecus ghari (um ca.- 2.5
Mio) (Siehe: [SWW13]:S.477) - Gattung: Paranthropus (Australopithecus)
(ab ca. -2.7 Mio) (Siehe: [WE17j]).
Kann möglicherweise auch dem
Australopithecus zugerechnet werden (Siehe:
[SWW13]:S.475).- Spezies/ Art: Paranthropus (Australopithecus)
aethiopicus (ab ca. -2.6 Mio bis ca. -2.3
Mio) (Siehe: [SWW13]:S.478) - Spezies/ Art: Paranthropus (Australopithecus)
boisei (ab ca. -2.3 Mio bis ca. -1.4 Mio)
(Siehe: [SWW13]:S.478). Mit dem Australopithecus
boisei starb der Australopithecus
vermutlich aus. - Spezies/ Art: Paranthropus (Australopithecus)
robustus (ab ca. -1.8 Mio bis ca. -1.5
Mio) (Siehe: [SWW13]:S.478)
- Spezies/ Art: Paranthropus (Australopithecus)
- Gattung: Homo (ab ca. -2.5/ -2.0 Mio).
Im allgemeinen ist es schwierig, sehr klare
Einteilungen bei den vielfältigen Funden
vorzunehmen. Deswegen gibt es bei der
Zuordnung der Funde zu bestimmten Mustern
unterschiedliche Hypothesen verschiedener
Forscher. Drei dieser Hypothesen seien hier
explizit genannt:- Kontinuitäts-Hypothese: In dieser Hypothese
wird angenommen, dass es vom
homo ergaster aus viele unterschiedliche
Entwicklungszweige gegeben hat, die
aber letztlich alle zum homo sapiens
geführt haben. Die Vielfalt der Formen
in den Funden reflektiert also so eine
genetische Variabilität. - Multiregionen-Hypothese: In dieser Hypothese
wird angenommen, dass sich –
ausgehend vom homo ergaster – regional
ganz unterschiedliche Formen ausgebildet
haben, die dann – bis auf den homo sapiens
mit der Zeit ausgestorben sind - Out-of-Africa Hypothese: Neben
früheren Auswanderungen aus Afrika
geht es in dieser Hypothese darum, dass
sich nach allen neuesten Untersuchungen
sagen lässt, dass alle heute lebenden
Menschen genetisch zurückgehen auf
den homo sapiens, der ca. um -100.000
Jahren von Afrika aus kommend nach und
nach alle Erdteile besiedelt hat (Siehe:
[SWW13]:S.488ff, 499).
Natürlich ist auch eine Kombination der ersten
beiden Hypothesen möglich (und wahrscheinlich),
da es zwischen den verschiedenen Formen
immer wieder Vermischungen geben
konnte. - Kontinuitäts-Hypothese: In dieser Hypothese
- Spezies/ Art: Homo rudolfensis (von
ca. -2.4 bis ca. -1.8 Mio) (Siehe:
[SWW13]:S.481) - Spezies/ Art: Homo habilis (von ca. -2.4 Mio bis ca. 1.65 Mio). Erste Art der Gattung Homo. Benutzte Steinwerkzeuge (Oldowan Kultur). Diese Artefakte sind
nachweisbar für -2.5 bis -700.000 (Siehe: [SWW13]:S.480) - Gattung: Australopithecus sediba (um ca.
-2 Mio) (Siehe: [SWW13]:S.477) - Spezies/ Art: Homo gautengensis (von ca.
-1.9 Mio bis ca. -0.6 Mio)(Südafrika) (Siehe:
[WE17h]) - Spezies/ Art: Homo ergaster (von ca. -1.9
Mio bis ca. -1.0 Mio) Werkzeuggebrauch
wahrscheinlich, ebenso die Nutzung von
Feuer (Lagerplätze mit Hinweisen um ca.
-1.6 Mio). Stellung zu homo erectus unklar.
(Siehe: [SWW13]:S.482f) Funde in
Nordafrika (ca. -1.8 Mio), Südspanien (ca. –
1.7-1.6 Mio und -1 Mio), Italien (ca. -1 Mio),
Israel (ca. -2 Mio), Georgien (ca. -1.8 bis –
1.7 Mio) und China (ca. -1.0 Mio) zeigen,
dass homo ergaster sich schon sehr früh
aus Afrika heraus bewegt hat. - Spezies/ Art: Homo erectus (Siehe:
[WE17f]) (ab ca. -1.9 Mio bis ca. -85.000/
-56.000); entwickelte sich vor allem in
Asien (China, Java…), möglicherweise
hervorgegangen aus dem homo ergaster.
Ist fas zeitgleich zu homo ergaster in Afrika
nachweisbar. Würde voraussetzen, dass
homo ergaster in ca. 15.000 Jahren den
Weg von Afrika nach Asien gefunden hat.
(Siehe: [SWW13]:S.484-487) - Spezies/ Art: Homo antecessor (Siehe:
[WE17e]) (von ca. -1.2 Mio bis –
800.000). Hauptsächlich Funde in
Nordafrika und Südspanien. Wird zur
ersten Auswanderungswelle ’Out of Africa’
gerechnet, die nach Europa und Asien kam.
Letzte Klarheit fehlt noch. Es scheint viele
Wechselwirkungen zwischen h.ergaster,
h.erectus, h.antecessor, h.heidelbergensis,
h.rhodesiensis, h.neanderthalensis sowie
h.sapiens gegeben zu haben. (Siehe:
[SWW13]:S.489) - Spezies/ Art ?: Homo cepranensis
(Datierung zwischen ca. -880.000 bis
ca.-440.000); (Siehe: [WD17k]) noch keine
klare Einordnung (siehe Anmerkungen zu
h.antecessor.) - Spezies/ Art: Homo heidelbergensis
(Siehe: [WD17l]) (von ca. -600.000 bis
-200.000). Überwiegend in Europa; es
gibt viele Ähnlichkeiten mit Funden
außerhalb von Europa in Afrika, Indien,
China und Indonesien, z.B. Ähnlichkeiten
zu homo rhodesiensis. Steinwerkzeuge,
weit entwickelte Speere, Rundbauten,
Feuerstellen, evtl. auch Kultstätten. (Siehe:
[SWW13]:SS.490-493). - Spezies/ Art: Homo rhodesiensis (Siehe:
[WE17i]) (von ca.-300.000 bis ca. –
125.000)(Ost- und Nord-Afrika, speziell
Zambia) - Spezies/ Art: Homo neanderthalensis
(ab ca. -250.000 bis ca. -33.000). Frühe
Formen und späte Formen. Genetische
Eigenentwicklung seit ca. -350.000/ -400.000. Schwerpunkt Europa, aber Ausdehnung von Portugal, Spanien, bis
Wales, Frankreich, England, Deutschland,
Kroatien, schwarzes Meer, Nordirak,
Zentralasien, Syrien, Israel . Meist nie
mehr als insgesamt einige 10.000 in ganz
Europa. In der Schlussphase parallel
mit homo sapiens für ca. 50.000 Jahre.
Es kam zu geringfügigen genetischen
Beeinflussungen. Eine eigenständige
hohe Werkzeugkultur, die aus der
Steinwerkzeugkultur der Acheul´een ca.
-200.000 hervorging und bis -40.000
nachweisbar ist. Neben Steinwerkzeugen
auch Schmuck. Sie pflegten Kranke,
bestatteten ihre Toten. Die differenzierte
Sozialstruktur, das gemeinsames Jagen,die
Werkzeugkultur, das großes Gehirn
sowie die Genbeschaffenheit lassen es
wahrscheinlich erscheinen, dass der
Neandertalerüber Sprache verfügte. Ein
besonders kalter Klimaschub um -50.000
verursachte einen starken Rückzug aus
West- und Mitteleuropa, der dann wieder
mit Einwanderer aus dem Osten gefüllt
wurde. Im Bereich Israels/ Palästina gab
es zwischen ca. -120.000 und -50.000
eine Koexistenz von Neandertaler und
homo sapiens. Was auch darauf hindeutet,
dass eine erste Auswanderungswelle von
h.sapiens schon um ca. -120.000/ -100.000
stattgefunden hatte, aber nur bis Israel
gekommen ist. Warum die Neandertaler
ausstarben ist unbekannt. homo sapiens
hat seine Population in Europa im Zeitraum
-55.000 und -35.000 etwa verzehnfacht.
(Siehe: [SWW13]:SS.493-498) - Spezies/ Art: Homo sapiens (ab ca. -190.000 bis heute); Wanderungsbewegungen aus Afrika heraus ab ca. -125.000
erster Vorstoß bis Arabien. Parallel gab
es eine kleine Auswanderung um -120.000
über das Niltal bis Palästina/Israel, die
aber keine weitere Expansion zeigte. Um
-70.000 von Arabien aus in den Süden des mittleren Ostens, um ca. -60.000/ -50.000 nach Neuguinea und
Australien. Vor ca. -50.000 bis -45.000 über
Kleinasien nach Südost-, Süd- und Westeuropa.
Um ca. -40.000 über Zentralasien
bis Nordchina. Von Asien aus um -19.000/ -15.000 Einwanderung in Nordamerika über
die Beringstraße, bis nach Südamerika um
ca. -13.000. Es gibt aber auch die Hypothese,
dass Südamerika schon früher
(ca. -35.000 ?)über den Pazifik besiedelt
wurde. Die Gene der Indianer in Nord- und
Südamerika stimmen mit Menschen aus
Sibirien, Nordasien und Südasien überein.
Ab der Zeit -60.000/ -40.000 wird ein deutlicher
kultureller Entwicklungssprung beim
homo sapiens diagnostiziert, dessen Entwicklung
seitdem anhält und sich heute
noch erheblich beschleunigt. Felszeichnungen
ab ca. -40.000, Werkzeuge, Wohnungen,
Kleidung, Sprache. - Spezies/ Art: Homo floresiensis
(Siehe: [WE17g])(ca. um -38.000 bis -12.000)(Insel Flores, Indonesien). Benutze Steinwerkzeuge, beherrschte das Feuer, Kleinwüchsig, entwickeltes Gehirn. Insel
war seit mindestens -800.000 besiedelt.
Vorfahren könnten seit -1 Mio dort gewesen
sein. (Siehe: [SWW13]:S.487f) - Spezies/ Art: Denisovaner (noch kein
wissenschaftlicher Name vereinbart)(um
-40.000) (Siehe: [WD17c]), Funde im
Altai Gebirge (Süd-Sibierien); es gibt
Funde auf den Pilippinen, in Indonesien,
Neuguinea, Australien, auf einigen Inseln
des südlichen Pazifik, mit den Genen der
Denisovaner. Herkunft möglicherweise von
h.heidelbergensis. Es gab genetischen
Austausch mit h.sapiens. (Siehe:
[SWW13]:S.498)
- Gattung: Orrorin tugenensis (ab ca. -6.2 bis
- Unterfamilie: Homininae
- Familie: Hominidae (Menschenaffen)(Aufteilung ab
- Überfamilie: Hominoidea (Menschenartige)
VIII. WAS FOLGT AUS ALLEDEM?
Jeder, der diesen Text bis hierher gelesen haben
sollte, wird sich unwillkürlich fragen: Ja und, was heißt
das jetzt? Was folgt aus Alledem?
In der Tat ist dieser Text noch nicht abgeschlossen.
Der Text stellt allerdings eine notwendige
Vorüberlegung dar zu der – hoffentlich – weiter führenden
Frage nach der Besonderheit des homo sapiens als
Erfinder und Nutzer von intelligenten Maschinen.
Während die abschließende Definition von potentiell
intelligenten Maschinen mit dem mathematischen
Konzept der Turingmaschine im Prinzip vollständig
vorliegt, erscheint die Frage, wer oder was denn der
homo sapiens ist, je länger umso weniger klar. Mit
jedem Jahr empirischer Forschung (in allen Bereichen)
enthüllt sich scheibchenweise eine immer unfassbarere
Komplexität vor unseren Augen, die ein Verständnis
des homo sapiens samt seinem gesamten biologischen
Kontext eher in immer weitere Ferne zu rücken scheint.
Konnten die großen Offenbarungsreligionen über
viele Jahrhunderte irgendwie glauben, dass sie
eigentlich wissen, wer der Mensch ist (obwohl sie
nahezu nichts wussten), ist uns dies heute – wenn wir
die Wissenschaften ernst nehmen – immer weniger
möglich. Wenn im jüdisch-christlichen Glauben der
Mensch bildhaft als ’Ebenbild Gottes’ bezeichnet werden
konnte und damit – zumindest indirekt – angesichts
dieser unfassbaren Erhabenheit eine Art Schauer über
den Rücken jagen konnte (ohne dass zu dieser Zeit
verstehbar war, worin denn die Besonderheit genau
besteht), so werden wir in den letzten Jahren durch
immer tiefere Einblicke in die Abgründe der Komplexität
von Leben und Lebensprozessen in einem scheinbar
lebensfremden physikalischen Universum provoziert,
herausgefordert, und Gelegenheit zum Staunen gäbe es
allerdings genug.
In diesem anwachsenden Wissen um
unser Nichtwissen begegnen wir einer schwer fassbaren
Größe, die wir salopp ’biologisches Leben’ nennen, die
aber alles übersteigt, dessen wir denkerisch fähig sind.
Eine der vielen Paradoxien des Universums ist
genau dieses Faktum: in einem scheinbar ’leblosen’
physikalischen Universum ’zeigen sich’ materielle
Strukturen, die Eigenschaften besitzen, die es strikt
physikalisch eigentlich nicht geben dürfte, und die sich
in einer Weise verhalten, die das ganze Universum
prinzipiell zu einem ’Un-Ort’ machen: das bekannte
physikalische Universum ist lebensfeindlich, das
biologische Leben will aber genau das Gegenteil:
es will leben. Wie kann das zusammen gehen? Warum
kann ein scheinbar lebloses physikalisches Universum
Überhaupt der Ort sein, wo Leben entsteht, Leben
stattfinden will, Leben sich schrittweise dem inneren
Code des ganzen Universums bemächtigt?
In weiteren Beiträgen wird es darum gehen, dieses
Phänomen ’biologisches Leben’ weiter zu erhellen,
und zu zeigen, wie das biologische Leben sich mit
Hilfe intelligenter Maschinen nicht nur generell weiter
entwickeln wird, sondern diesen Prozess noch erheblich
beschleunigen kann. Es gilt hier die Arbeitshypothese,
dass die intelligenten Maschinen ein konsequentes
Produkt der biologischen Evolution sind und dass es
gerade dieser Kontext ist, der dieser Technologie ihre
eigentliche Zukunftsfähigkeit verleiht.
Die heutigen Tendenzen, die Technologie vom biologischen Leben
zu isolieren, sie in dieser Isolation zugleich in geradezu
religiöser Manier zu Überhöhen, wird die evolutionär
induzierte Entwicklung dieser Technologie eher
behindern, und damit auch das vitale Element der
biologischen Evolution, den homo sapiens.
Der homo sapiens ist kein Individuum, er wird
repräsentiert durch eine Population, die wiederum nur
Teil einer umfassenderen Population von Lebensformen
ist, die sich gegenseitig am und im Leben halten. Es wird
wichtig sein, dass der homo sapiens diese Arbeitsteilung
versteht, bevor er mit seiner wachsenden Aufbau- und
Zerstörungskraft das biologische Universum zu stark
beschädigt hat.
Zum aktuellen Zeitpunkt kann niemand mit Gewissheit
sagen, ob das alles irgendeinen ’Sinn’ besitzt, d.h. ob es
in all den Abläufen in der Zukunft eine Menge möglicher
Zielzustände gibt, die in irgendeinem Sinne als ’gut’/
’schön’/ ’erfüllend’ oder dergleichen bezeichnet werden
können. Genauso wenig kann aber irgend jemand zum
aktuellen Zeitpunkt mit Gewissheit einen solchen Sinn
ausschließen. Rein empirisch kann man schon heute
eine solche Menge an atemberaubenden Strukturen und
Zusammenhänge erfassen, die ’aus sich heraus’ ein
Exemplar der Gattung homo sapiens in ’Erstaunen’ und
’Begeisterung’ versetzen können; aber weder gibt es für
solch ein Erstaunen einen Zwang, eine Regel, ein Muss,
noch ist all dies ’zwingend’. Noch immer können wir
nicht ausschließen, dass dies alles nur ein Spiel ist,
eine gigantische kosmologische Gaukelei, oder – wie
es die physikalischen kosmologischen Theorien nahelegen
– in einem gigantischen Kollaps endet, aus der
möglicherweise wieder ein Universum entsteht, aber ein
anderes.
REFERENCES
- [AJL+15] B. Alberts, A. Johnson, J. Lewis, D. Morgan, M. Raff,
K. Roberts, and P. Walter.Molecular Biology of the Cell.
Garland Science, Taylor & Francis Group, LLC, Abington
(UK) – New York, 6 edition, 2015. - [GJ79] Michael R. Garey and David S. Johnson.Computers and
Intractibility. A Guide to the Theory of NP.Completeness.
W.H. Freeman an Company, San Francisco (US), 1 edition,
1979. - [Keg15] Bernhard Kegel.Die Herrscher der Welt. DuMont, Köln (DE),
1 edition, 2015. - [Kuh62] Thonas S. Kuhn.The Structure of Scientific Revolutions.
University of Chicago Press, Chicago (US), 1 edition, 1962. - [Par15] Hermann Parzinger.DIE KINDER DES PROMETHEUS.
Geschichte der Menschheit vor der Erfindung der Schrift.
Wissenschaftliche Buchgesellschaft, Darmstadt (DE), 2 edition,
2015. - [Rot00] Peter Rothe.Erdgeschichte. Spurensuche im Gestein. Wissenschaftliche
Buchgesellschaft, Darmstaadt (DE), 1 edition, 2000. - [SWW13] Volker Storch, Ulrich Welsch, and Michael Wink, editors.
Evolutionsbiologie. Springer-Verlag, Berlin – Heidelberg, 3 edition, 2013. - [WD17a] Wikipedia-DE. Archäologie. 2017.
- [WD17b] Wikipedia-DE. Ardipithecus. 2017.
- [WD17c] Wikipedia-DE. Denisova-mensch. 2017.
- [WD17d] Wikipedia-DE. Eiszeitalter. 2017.
- [WD17e] Wikipedia-DE. Evolutionsbiologie. 2017.
- [WD17f] Wikipedia-DE. Genetik. 2017.
- [WD17g] Wikipedia-DE. Geologie. 2017.
- [WD17h] Wikipedia-DE. Geschichte der geologie. 2017.
- [WD17i] Wikipedia-DE. Graecopithecus freybergi. 2017.
- [WD17j] Wikipedia-DE. Heisenbergsche unschärferelation. 2017.
- [WD17k] Wikipedia-DE. Homo cepranensis. 2017.
- [WD17l] Wikipedia-DE. Homo heidelbergensis. 2017.
- [WD17m] Wikipedia-DE. Kenyanthropus platyops. 2017.
- [WD17n] Wikipedia-DE. Klimatologie. 2017.
- [WD17o] Wikipedia-DE. Känozoisches eiszeitalter. 2017.
- [WD17p] Wikipedia-DE. Kosmologie. 2017.
- [WD17q] Wikipedia-DE. Kurt gödel. 2017.
- [WD17r] Wikipedia-DE. Menschenaffen. 2017.
- [WD17s] Wikipedia-DE. Molekularbiologie. 2017.
- [WD17t] Wikipedia-DE. Orrorin tugenensis. 2017.
- [WD17u] Wikipedia-DE. Paläontologie. 2017.
- [WD17v] Wikipedia-DE. Sahelanthropus tchadensis. 2017.
- [WD17w] Wikipedia-DE. Supervulkan. 2017.
- [WE17a] Wikipedia-EN. chimpanzee–human last common ancestor
(chlca). 2017. - [WE17b] Wikipedia-EN. Cosmology. 2017.
- [WE17c] Wikipedia-EN. Earth science. 2017.
- [WE17d] Wikipedia-EN. Homininae. 2017.
- [WE17e] Wikipedia-EN. Homo antecessor. 2017.
- [WE17f] Wikipedia-EN. Homo erectus. 2017.
- [WE17g] Wikipedia-EN. Homo floresiensis. 2017.
- [WE17h] Wikipedia-EN. Homo gautengensis. 2017.
- [WE17i] Wikipedia-EN. Homo rhodesiensis. 2017.
- [WE17j] Wikipedia-EN. Paranthropus. 2017.
- [WE17k] Wikipedia-EN. Sahelanthropus tchadensis. 2017.
- [WE17l] Wikipedia-EN. Simian. 2017.
Eine FORTSETZUNG findet sich HIER.
VIII. KONTEXTE
Einen Überblick über alle Blogeinträge von Autor cagent nach Titeln findet sich HIER.
Einen Überblick über alle Themenbereiche des Blogs findet sich HIER.
Das aktuelle Publikationsinteresse des Blogs findet sich HIER.